




Uno dei punti di forza della filiera bufalina è sicuramente la qualità della materia prima, il latte di bufala, dal quale si ricavano principalmente mozzarella ed in quantità minori, ma non trascurabili, altri derivati come la ricotta e lo yogurt. Oltre alla bontà indiscussa di questi prodotti, essi contengono molecole biologicamente attive che conferiscono alla mozzarella, alla ricotta, ed allo yogurt di bufala proprietà nutraceutiche; di conseguenza, questi prodotti, se inseriti in una dieta equilibrata, possono essere considerati “alimenti funzionali”. Studi recenti mostrano che la concentrazione di tali molecole può variare a seconda delle tecniche di allevamento adottate, evidenziando sia l’importanza di incentrare l’allevamento intorno al benessere animale, sia una possibile rivalutazione del ruolo del siero del latte.
In letteratura è noto che il latte di bufala è un alimento ricco in proteine ad elevato potere biologico. Queste, durante i processi di lavorazione lattiero-caseari o durante la digestione enzimatica, possono essere scisse in frammenti proteici, che prendono il nome di peptidi bioattivi i quali espletano diversi effetti benefici sull’organismo. In base alla posizione occupata nella catena madre ed alla proteina da cui derivano, le azioni possono essere molteplici. Negli ultimi anni nel latte è stata registrata la presenza, oltre dei peptidi bioattivi, anche di un’altra famiglia di molecole, i metaboliti bioattivi, che sono abbondanti nel latte dei ruminanti. Tra queste molecole, particolarmente interessanti sono i precursori della carnitina, come acetil-carnitina, propionil-carnitina e butirril-carnitina, e le betaine, come γ-butirrobetaina, glicinabetaina e δ-valerobetina. Le proprietà biologiche di tali molecole, ed in particolare della δ-valerobetina, sono molto interessanti in quanto svolgono un’attività antiossidante, antinfiammatoria ed antitumorale (D’Onofrio et al. 2020).
La concentrazione dei metaboliti nel latte è influenzata da molteplici fattori come specie, razza e tecniche di allevamento. In generale, si può affermare che la presenza di metaboliti bioattivi è maggiore nel latte dei ruminanti rispetto a quello dei non ruminanti (Servillo et al. 2018). Differenze statisticamente significative si sono inoltre registrate sia tra differenti razze appartenenti alla stessa specie, sia nell’ambito della stessa razza variando le tecniche di allevamento. In particolar modo, si è visto che l’alimentazione con foraggio verde e la disponibilità di spazio pro-capite possono avere influenza sulla qualità del prodotto finale (Salzano et al. 2019, 2020). È importante tenere presente che le caratteristiche qualitative ritrovate nel latte di bufala vanno ricercate e studiate soprattutto nei suoi derivati, in quanto sono i principali prodotti consumati.
Alla luce di queste considerazioni sono state effettuate due sperimentazioni volte a verificare gli effetti delle tecniche di allevamento alternative sulla produzione di biomolecole nel latte e derivati bufalini.
Esperimento 1 – Utilizzo di foraggio verde nella razione
Nel primo esperimento è stato inserito il foraggio verde nella razione delle bufale in ragione del 30%. Lo studio ha avuto una durata di due mesi ed è stato condotto presso un allevamento bufalino situato nel sud Italia. Le bufale in prova sono state equamente divise in due gruppi sperimentali, con razioni isoproteiche e isoenergetiche che differivano solo per l’inclusione (Gruppo T) o meno (Gruppo C) del 30% di foraggio di erba medica. La scelta del 30% non è casuale, ma deriva da precedenti dati in letteratura (Pastorini et al 2019; Morales-Almaráz et al., 2010) che mostravano come l’aumento di foraggio verde nella razione, fino ad un massimo del 30%, non andava ad influenzare l’ingestione di sostanza secca degli animali.
Esperimento 2 – Disponibilità di spazio pro-capite
Durante il secondo studio, invece, è stato valutato l’effetto della differenza di spazio-procapite. Il lavoro è stato eseguito su 96 bufale che sono state divise in due gruppi. Anche in questo caso lo studio è durato due mesi. Nel gruppo S10 le bufale avevano a disposizione 10 m2 pro-capite, mentre nel Gruppo S15 avevano 15 m2 pro-capite. Anche in questo caso la scelta della disponibilità di spazio deriva da dati precedenti in letteratura che mostravano come fino ad uno spazio minimo di 10 m2 capo gli animali potevano essere allevati senza ledere la sfera produttiva e riproduttiva (Salzano et al., 2018).
Per entrambe le sperimentazioni, le bufale sono state assegnate ai due gruppi in base al peso, agli anni, ai giorni di lattazione, all’ordine di parto ed alla produzione di latte. Le bufale sono state allevate in paddock e munte due volte al giorno. Dopo un periodo di adattamento alle nuove condizioni di 14 giorni, nel primo esperimento sono stati prelevati ed analizzati due campioni al mese di latte, siero, ricotta, yogurt e mozzarella. Nel secondo esperimento, invece, sono stati prelevati e analizzati due campioni al mese di latte, siero, mozzarella e ricotta. Il dosaggio delle carnitine e delle betaine è stato eseguito tramite analisi HPLC-ESI-MS\MS, mentre per confrontare i dati è stata utilizzata l’analisi della varianza ad una via (ANOVA) ed i dati sono riportati come media di gruppo con relativa deviazione standard (DS).
Risultati Esperimento 1 – Utilizzo di foraggio verde nella razione
Nella tabella 1 sono mostrati i risultati riguardanti l’inclusione del 30% di foraggio verde (erba medica) nella razione. Le analisi hanno evidenziato un significativo incremento di carnitina, acetilcarnitina, propionilcarnitina e δ-valerobetaina (P<0.05) nel gruppo C rispetto al gruppo T nelle diverse matrici alimentari. Questo risultato è dovuto all’abbondanza di Trimetilisina (TML) nell’erba medica, e più in generale nei foraggi verdi a foglia larga che ne contengono quantità relativamente elevate. La TML rappresenta il substrato iniziale della sintesi endogena della carnitina e dei suoi precursori. A differenza del gruppo T, al gruppo C è stata somministrata una razione totalmente miscelata, TMR (total mixed ratio) o unifeed, che rappresenta una scarsa fonte di TML in quanto ricca in glutine di frumento o proteine di soia il cui contenuto in TML è trascurabile (Servillo et al., 2018).
Tabella 1 – Livelli di biomolecole funzionali (mg/L) in latte, yogurt, mozzarella, ricotta, siero registrati nei due differenti gruppi.
| Biomolecole2 espresse in mg/l |
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Prodotto | Gruppo1 | Car | C2Car | C3Car | IC4Car | nC4car | δ-VB | γ-BB | glyBet |
| Latte | GC | 31.5±0.8a | 39.1±1.3a | 13.5±1.3a | 18.3±0.4a | 4.8±0.3 | 7.01±1.7 | ||
| GT | 41.6±0.5b | 49.7±0.8b | 20.5±1.7b | 21.8±0.4b | 3.3±1.1 | 7.11±0.7 | |||
| Yogurt | GC | 35.7±1.7 | 24.8±5.2 | 25.5±5.6 | 10.9±0.5 | 11.6±0.8 | 17.2±0.5 | 5.6±0.2 | 5.2±0.5 |
| GT | 31.1±3.6 | 20.6±2.0 | 23.0±1.6 | 8.6±1.2 | 8.4±0.6 | 14.0±3.0 | 3.4±0.6 | 4.9±0.2 | |
| Mozzarella | GC | 15.1±1.2 | 8.8±0.3 | 1.4±0.1 | 0.6±0.1 | 0.4±0.1 | 1.6±0.3 | 6.3±0.1 | 2.0±0.2 |
| GT | 13.6±0.1 | 7.3±1.2 | 1.1±0.2 | 0.6±0.1 | 0.4±0.1 | 1.4±0.2 | 5.0±0.8 | 2.0±0.7 | |
| Ricotta | GC | 23.8±0.3 | 15.9±0.4 | 7.2±1.1 | 3.3±0.1 | 2.2±0.2 | 1.2±0.2 | 13.2±0.8 | 6.6±0.7 |
| GT | 21.1±0.5 | 13.8±0.6 | 5.9±1.0 | 3.0±0.5 | 1.9±0.3 | 1.4±0.4 | 11.0±2.3 | 14.9±1.3 | |
| Siero | GC | 39.2±0.1 | 24.1±2.5 | 27.1±2.5 | 11.4±0.5 | 15.8±0.5 | 2.2±0.3 | 23.3±0.8 | 9.3±0.3 |
| GT | 31.2±0.5 | 24.4±2.5 | 25.1±0.9 | 10.6±0.0 | 14.7±0.5 | 2.7±0.3 | 20.9±0.5 | 8.5±0.7 | |
2Car = l-carnitina; C2Car = acetilcarnitina; C3Car = propionilcarnitina; δ-VB = δ-valerobetaina; γ-BB = γ-butirrobetaina; iC4Car = isobutirrilcarnitina; nC4Car = butirrilcarnitina; glyBet = glicina betaina.1 GC= gruppo controllo GT= gruppo trattatoab, I valori all’interno delle colonne con apici diversi differiscono; P <0,05
È stato interessante rappresentare, anche, graficamente la concentrazione di carnitina e δ-valerobetaina dei due gruppi nel latte e nei suoi derivati.
Figura 1 – Confronto nei due gruppi dei livelli di carnitina nel latte e nei suoi derivati.
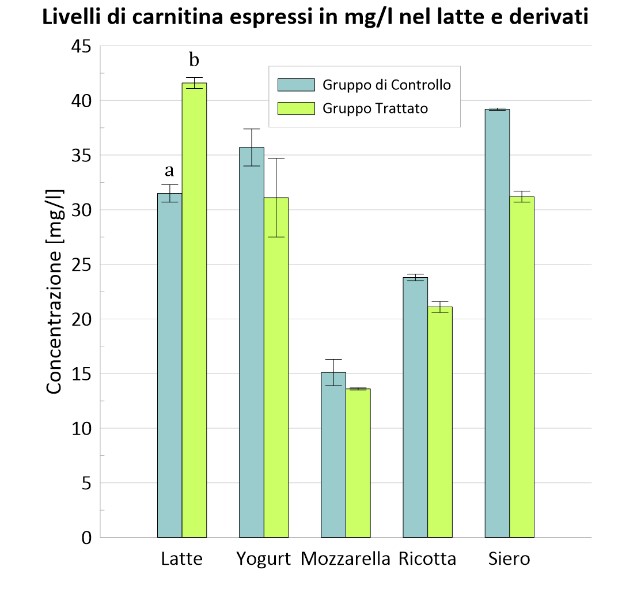
a, b, I valori all’interno delle colonne con apici diversi differiscono; P <0,01.
Figura 2 – Confronto nei due gruppi dei livelli di δ-VB nel latte e nei suoi derivati.
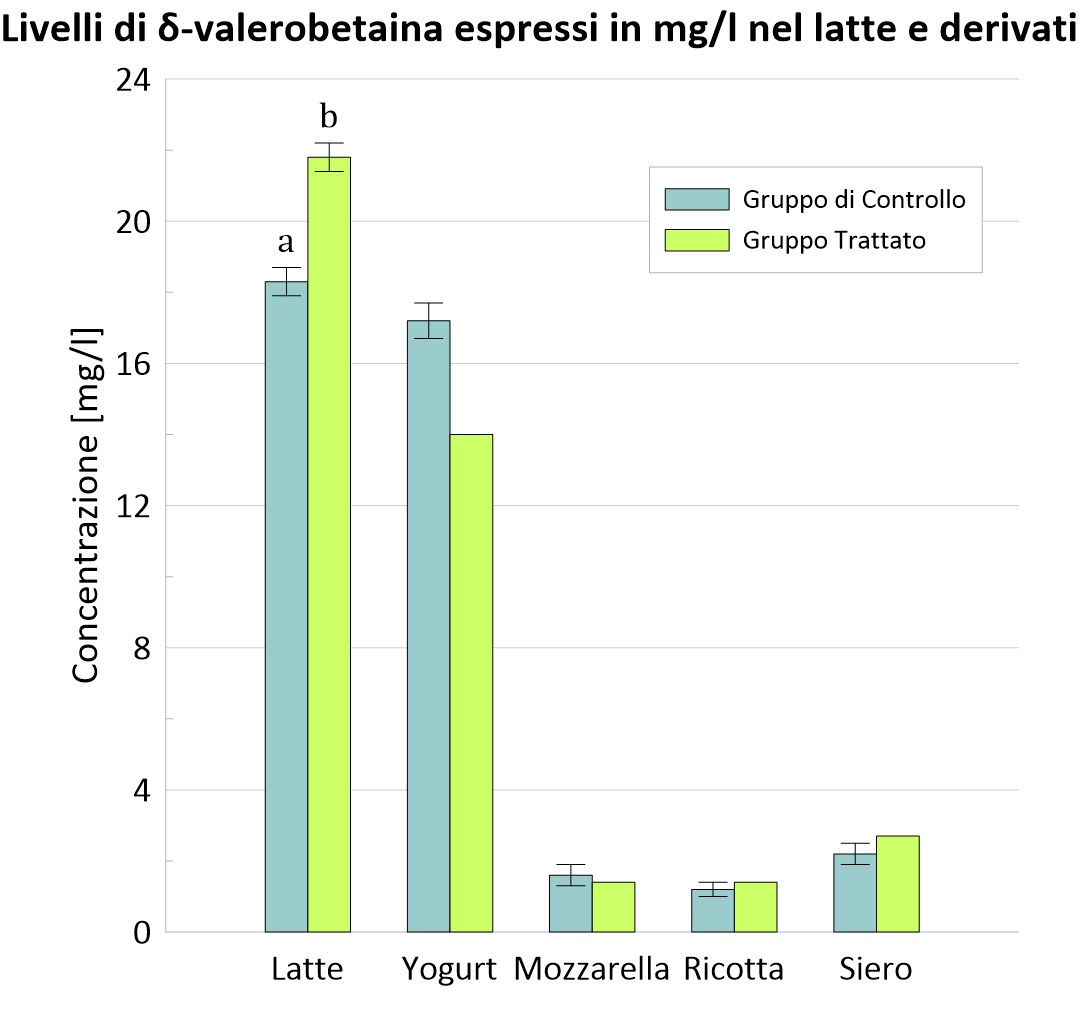
a, b, I valori all’interno delle colonne con apici diversi differiscono; P <0,05.
Risultati Esperimento 2 – Disponibilità di spazio pro-capite
Come per i risultati precedenti è riportata la tabella riassuntiva dei valori registrati. L’incremento di carnitina e δ-valerobetaina nel latte di bufala e nel siero di latte è dovuto probabilmente al fatto che animali allevati con minor spazio procapite sono più stressati. Quando l’animale si trova in questa condizione, attraverso l’attivazione dell’asse ipotalamo-ipofisi-surrene, vengono secreti i glucorticoidi. Questi ormoni comportano la riduzione del consumo energetico in vari distretti dell’organismo per favorirne l’approvvigionamento dell’apparato locomotore e del SNC (sistema nervoso centrale), e più in generale una maggiore capacità d’utilizzo dei diversi substrati metabolici, con attivazione dei processi di gluconeogenesi. Nel gruppo S15, invece, tale condizione non si è manifestata e lo spostamento del rapporto glicolisi/gluconeogenesi a favore della glicolisi potrebbe giustificare l’abbondanza della carnitina e dei suoi precursori nel latte e derivati bufalini (Salzano et al. 2019).
Tabella 2 – Livelli di biomolecole funzionali (mg/L) in latte, siero, mozzarella, ricotta registrati nei due differenti gruppi.
| Biomolecole2 espresse in mg/l | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Prodotto | Gruppo1 | Car | C2Car | C3Car | IC4Car | nC4car | γ-BB | δ-VB | glyBet |
| Latte | S15 | 56,7±1,1A | 51,9±0,3A | 34,8±1,0A | 10,9±0,6 | 14,2±1,2 | 6,6±0,2 | 24,2±0,5A | 23,1±2,0A |
| S10 | 39,8±0,7B | 39,7±0,7B | 21,0±0,9B | 11,3±0,9 | 12,6±0,9 | 6,1±0,2 | 16,7±0,5B | 13,5±1,6B | |
| Siero | S15 | 40,9±0,8A | 41,1±1,7A | 26,9±0,8A | 10,4±1,2 | 14,3±1,2 | 6,0±0,4 | 22,0±0,9A | 10,7±0,4A |
| S10 | 31,7±0,7B | 28,7±2,6B | 17,6±1,2B | 11,3±1,3 | 13,6±1,3 | 5.9±0,3 | 15,5±0,7B | 7,9±0,5B | |
| Mozzarella | S15 | 27,0±0,7 | 12,2±1,0 | 6,0±0,3 | 2,4±0,2 | 2,6±0,1 | 2,3±0,2 | 6,1±0,1 | 4.3±0.5 |
| S10 | 28,3±1,3 | 12,0±1,0 | 6,3±0,3 | 2,1±0,2 | 2,5±0,2 | 2,6±0,3 | 5,9±0,1 | 3,8±0,4 | |
| Ricotta | S15 | 44,2±2,2 | 39,3±1,4 | 22,5±0,9 | 9,8±0,7 | 11,1±0,5 | 6,5±0,3 | 15,7±0,3 | 14,9±0,8 |
| S10 | 41,4±0,9 | 36,8±0,8 | 23,9±0,7 | 10,7±0,5 | 11,7±0,7 | 6,1±0,2 | 16,0±0,4 | 15,6±0,7 | |
2Car = l-carnitina; C2Car = acetilcarnitina; C3Car = propionilcarnitina; δ-VB = δ-valerobetaina; γ-BB = γ-butirrobetaina; iC4Car = isobutirrilcarnitina; nC4Car = butirrilcarnitina; glyBet = glicina betaina. 1 S15=spazio pro-capite di 15m2 S10= spazio pro-capite di 10 m2
AB, I valori all’interno delle colonne con apici diversi differiscono; P <0,01
Anche qui la concentrazione di carnitina e della δ-valerobetaina dei due gruppi nel latte e nei suoi derivati è stata riportata graficamente.
Figura 3 – Confronto tra i due gruppi dei livelli di carnitina nel latte e nei suoi derivati.
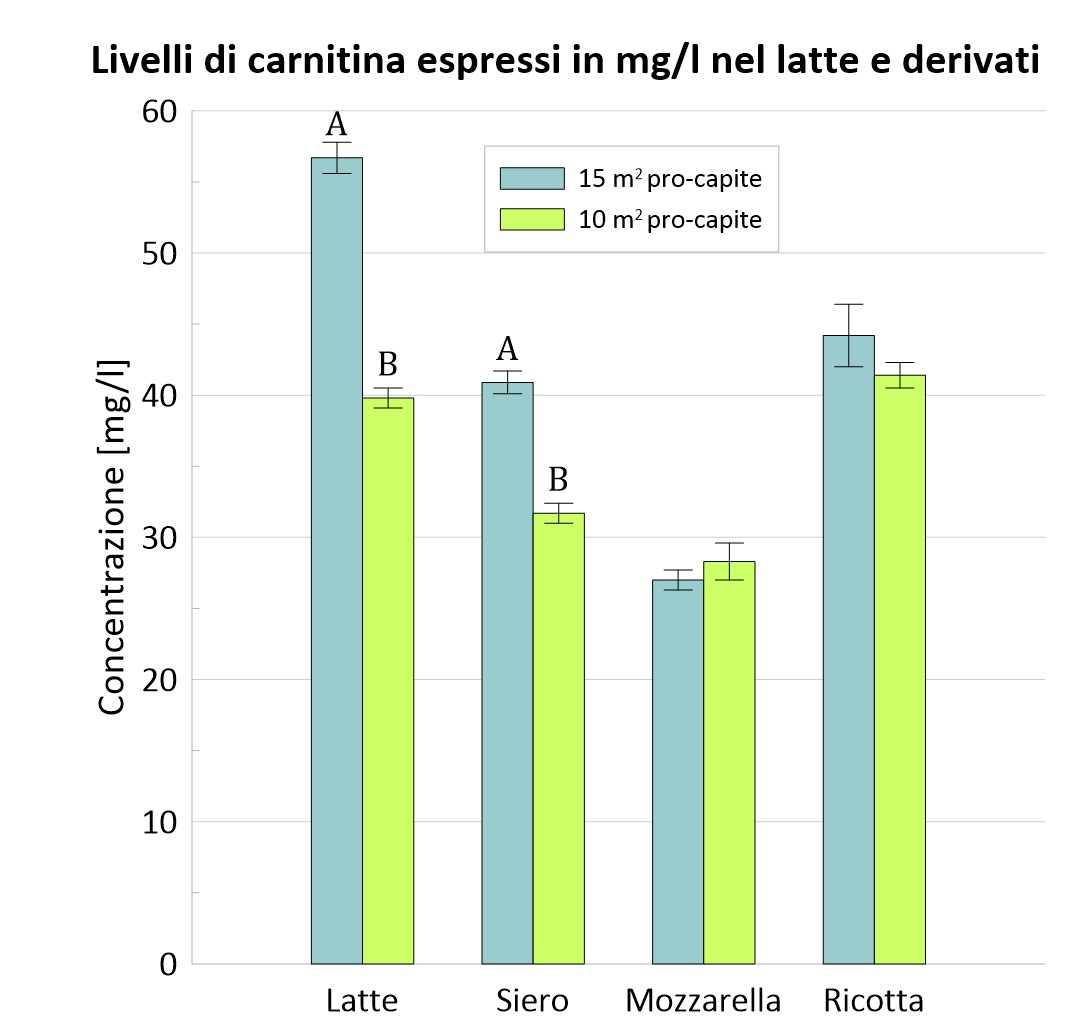
A, B, I valori all’interno delle colonne con apici diversi differiscono; P <0,01.
Figura 4 – Confronto tra i due gruppi dei livelli di carnitina nel latte e nei suoi derivati.
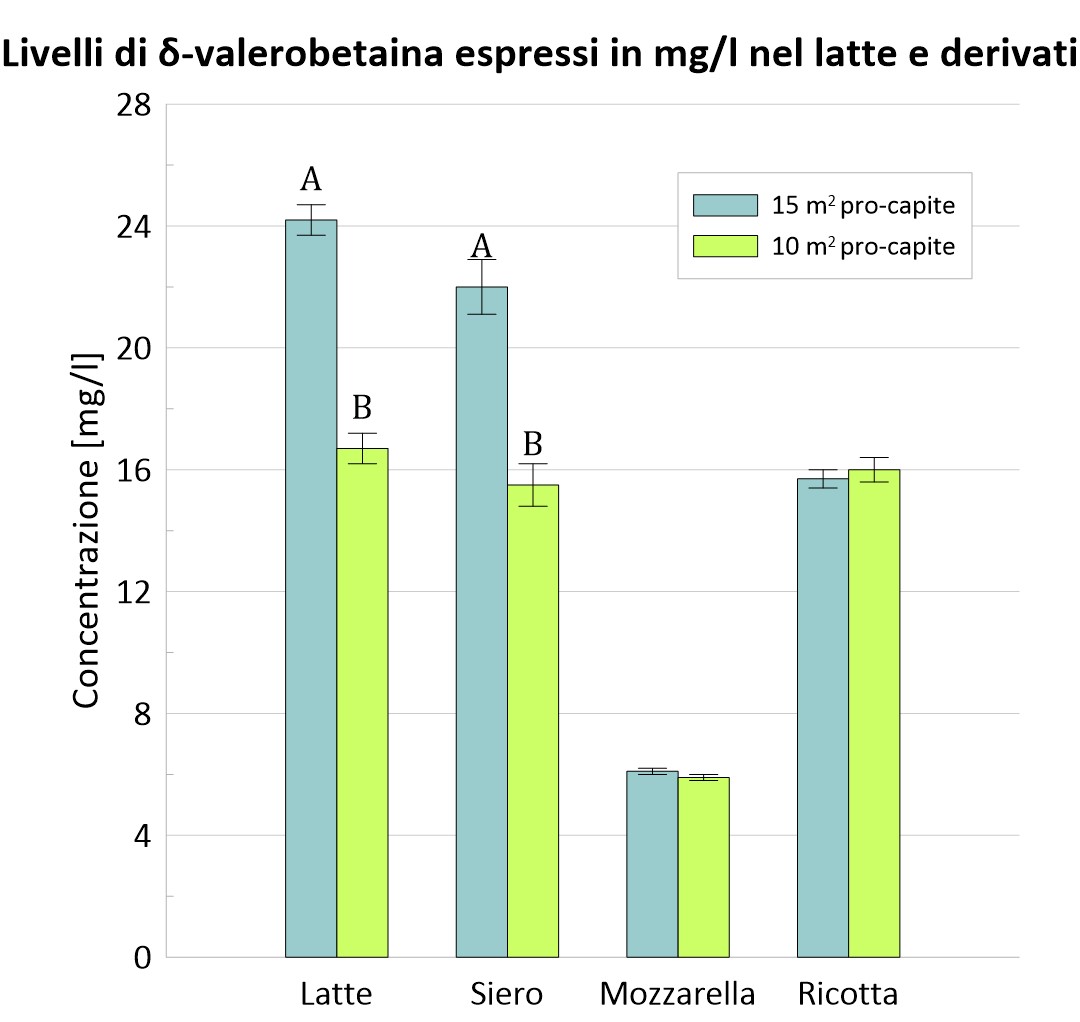
A, B, I valori all’interno delle colonne con apici diversi differiscono; P <0,01.
I risultati ottenuti evidenziano due importanti aspetti. In primo luogo, confermano che scegliere tecniche di allevamento che puntino all’“animal welfare” è fondamentale per incrementare i livelli di biomolecole nel latte e derivati. Assicurare il benessere animale nei sistemi di produzione zootecnici intensivi garantisce produttività, qualità e sicurezza igienico-sanitaria delle produzioni (Broom, 2010). L’esito delle prove ci indirizza verso una linea di pensiero che vede salute umana, ambientale e benessere animale legate tra di loro: questo concetto viene racchiuso nel termine “One Health”. Altro aspetto evidenziato è il ruolo del siero di latte. Quest’ultimo, insieme al latte di bufala, ha registrato le concentrazioni maggiori di biomolecole. Questa peculiarità è dovuta al fatto che i metaboliti bioattivi hanno dimensioni inferiori a quelle delle maglie caseiniche che si formano durante la cagliata, per cui non vengono trattenuti all’interno della mozzarella e si ritrovano perlopiù nel siero. Il siero di latte, da sempre, viene considerato l’inquinante più importante dell’industria lattiero-casearia, non solo per l’alto carico organico ma anche per i grandi volumi prodotti (Walzem et al., 2002). Tuttavia, la percezione del siero di latte come inquinante è cambiata con la scoperta delle sue proprietà funzionali e bioattive, diventando un prodotto aggiuntivo alla produzione di formaggio (De Boer, 2014). È proprio in quest’ottica che sembra essere ragionevole la possibilità di ampliare l’utilizzo di metaboliti funzionali presenti nel siero di latte, non solo nell’industria alimentare, ma anche come integratori naturali o di sintesi in industrie farmaceutiche, sanitarie e diagnostiche (FAO, 2011).
In definitiva, il latte di bufala ed i suoi derivati rappresentano un ottimo esempio di alimenti funzionali in quanto ricchi di molecole capaci di influenzare diversi meccanismi fisiologici nell’organismo umano. L’integrazione di latticini di bufala nella dieta, quindi, è sicuramente una mossa vincente per la tutela e la prevenzione della salute dei consumatori. A questo proposito, la zootecnia italiana, da sempre sinonimo di qualità, deve puntare sulla valorizzazione delle proprie produzioni riducendo al minimo gli sprechi. Questo settore è diretto responsabile dell’alimentazione umana per cui la qualità, intesa in ogni sua sfaccettatura (da quella organolettica a quella di igiene e sicurezza), è fondamentale per assicurare la salute dei consumatori e la sopravvivenza del settore. In futuro potrebbe essere interessante indagare sul comportamento di tali molecole nelle diverse fasi fisiologiche dell’animale e comprendere se la produzione di metaboliti è conseguenza di fattori genetici.
Sinossi tratta da: Short commu-nication: Space allocation in intensive Mediterranean buffalo production influences the profile of functional biomolecules in milk and dairy products – Salzano, A.; Licitra, F.; D’Onofrio, N.; Balestrieri, M.L.; Limone, A.; Campanile, G.; D’Occhio, M.J.; Neglia, G. – J Dairy Sci 2019, 102, 7717–22. doi: 10.3168/jds.2019-16656.
Green feed increases antioxidant and antineoplastic activity of buffalo milk: a globally significant livestock. – Salzano, A.; Neglia, G.; D’Onofrio, N.; Balestrieri, M.L.; Limone, A.; Cotticelli, A.; Marrone, R.; Anastasio, A.; D’Occhio, M.J.; Campanile, G. – Food Chem 2021, 344, 128669. doi: 10.1016/j.foodchem.2020.
Autori
Lucia Trapanese e Angela Salzano – Dipartimento di Medicina Veterinaria e Produzioni Animali, Università di Napoli “Federico II”.
Bibliografia
- Broom, Donald M. 2010. «Animal welfare: an aspect of care, sustainability, and food quality required by the public». Journal of veterinary medical education 37(1):83–88.
- De Boer, Ruud. 2014. From milk by-products to milk ingredients: upgrading the cycle. John Wiley & Sons.
- D’Onofrio, Nunzia, Luigi Mele, Elisa Martino, Angela Salzano, Brunella Restucci, Domenico Cautela, Marco Tatullo, Maria Luisa Balestrieri, e Giuseppe Campanile. 2020. «Synergistic Effect of Dietary Betaines on SIRT1-Mediated Apoptosis in Human Oral Squamous Cell Carcinoma Cal 27». Cancers 12(9):2468.
- Morales-Almaráz, E., Soldado, A., González, A., Martínez-Fernández, A., Domínguez-Vara, I., de la Roza-Delgado, B., &Vicente, F. (2010). Improving the fatty acid profile of dairy cow milk by combining grazing with feeding of total mixed ration. Journal of Dairy Research,77, 225–230.
- Pastorini M, Pomiés N, Repetto JL, Mendoza A, Cajarville C. Productive performance and digestive response of dairy cows fed different diets combining a total mixed ration and fresh forage. J Dairy Sci. 2019 May;102(5):4118-4130.
- Salzano Angela, Spagnuolo Maria Stefania, Lombardi Pietro, Vecchio Domenico, Limone Antonio, Bolletti Censi Sergio, Balestrieri Anna, Pelagalli Alessandra, Neglia Gianluca. Influences of different space allowance on reproductive performances in buffalo. Animal Reproduction vol. 14, nr. 2, April/June.
- Salzano Angela, Francesca Licitra, Nunzia D’Onofrio, Maria Luisa Balestrieri, Antonio Limone, Giuseppe Campanile, Michael J. D’Occhio, e Gianluca Neglia. 2019. «Short Communication: Space Allocation in Intensive Mediterranean Buffalo Production Influences the Profile of Functional Biomolecules in Milk and Dairy Products». Journal of Dairy Science 102(9):7717–22.
- Salzano Angela, Gianluca Neglia, Nunzia D’Onofrio, Maria Luisa Balestrieri, Antonio Limone, Alessio Cotticelli, Raffaele Marrone, Aniello Anastasio, Michael J. D’Occhio, e Giuseppe Campanile. 2020. «Green Feed Increases Antioxidant and Antineoplastic Activity of Buffalo Milk: A Globally Significant Livestock». Food Chemistry 344:128669.
- Servillo, Luigi, Nunzia D’Onofrio, Alfonso Giovane, Rosario Casale, Domenico Cautela, Domenico Castaldo, Francesco Iannaccone, Gianluca Neglia, Giuseppe Campanile, e Maria Luisa Balestrieri. 2018. «Ruminant meat and milk contain δ-valerobetaine, another precursor of trimethylamine N-oxide (TMAO) like γ-butyrobetaine». Food chemistry 260:193–99.
- Walzem, RL, CJ Dillard, e J. Bruce German. 2002. «Whey components: millennia of evolution create functionalities for mammalian nutrition: what we know and what we may be overlooking». Critical reviews in food science and nutrition 42(4):353–75.




